


| name | Amanita parvipantherina |
| name status | nomen acceptum |
| author | Zhu L. Yang, M. Weiss & Oberw. |
| english name | "Asian Small Panther Amanita" |
| images |
 1. Amanita parvipantherina, China. , Kunming, Yunnan Prov., China") 2. Amanita parvipantherina, Heilongtan (Black Dragon Pool), Kunming, Yunnan Prov., China |
| cap |
The cap of Amanita parvipantherina is 35 - 60 mm wide, convex to plano-convex, nonappendiculate, and having a tuberculate-striate margin. The cap is grayish to ochraceous, becoming brown to brownish over disc. The flesh is white. The volva is present in conical, subconical to granular, dirty white to grayish to yellowish remnants. |
| gills |
The gills are free, white, and crowded. The short gills are truncate. |
| stem |
The stem is 40 - 90 × 5 - 10 mm, subcylindric, and white. The stem bears a weakly structured pallid annulus. The volva is distributed over the stem in granular, dirty white to yellowish to grayish remnants. |
| spores |
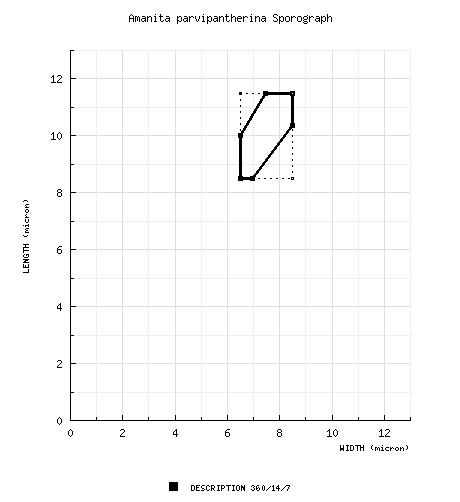
The spores measure 8.5 -11.5 × 6.5 - 8.5 µm and are broadly ellipsoid to ellipsoid and inamyloid. Clamps are not present at the bases of basidia. |
| discussion |
The species occurs on soil in mixed forests with Pine (Pinus). It fruits from July to August in Yunnan Prov., southwestern China, at 1200-2500 m elev. For species to compare within the "A. pantherina group," see the discussion of A. pantherina (DC. : Fr.) Krombh.—Zhu L. Yang |
| brief editors | RET |
| name | Amanita parvipantherina | ||||||||||||||||||||||||||||
| author | Zhu L. Yang, M. Weiss & Oberw. 2004. Mycologia 96: 643. | ||||||||||||||||||||||||||||
| name status | nomen acceptum | ||||||||||||||||||||||||||||
| english name | "Asian Small Panther Amanita" | ||||||||||||||||||||||||||||
| MycoBank nos. | 488566 | ||||||||||||||||||||||||||||
| GenBank nos. |
Due to delays in data processing at GenBank, some accession numbers may lead to unreleased (pending) pages.
These pages will eventually be made live, so try again later.
| ||||||||||||||||||||||||||||
| holotypes | HKAS 32492 | ||||||||||||||||||||||||||||
| intro |
The following text may make multiple use of each data field. The field may contain magenta text presenting data from a type study and/or revision of other original material cited in the protolog of the present taxon. Macroscopic descriptions in magenta are a combination of data from the protolog and additional observations made on the exiccata during revision of the cited original material. The same field may also contain black text, which is data from a revision of the present taxon (including non-type material and/or material not cited in the protolog). Paragraphs of black text will be labeled if further subdivision of this text is appropriate. Olive text indicates a specimen that has not been thoroughly examined (for example, for microscopic details) and marks other places in the text where data is missing or uncertain. The following material is derived entirely from the protolog of the present taxon. NOTE: Spore data from papers by Z. L. Yang are presented following his use of the "Times New Roman" face for "Q" and "Q'"—respectively, " | ||||||||||||||||||||||||||||
| pileus | from protolog: 35 - 60 mm wide, grayish to ocrhraceous (Light Drab, Buffy Brown, 5B2-3, 5C4-6), becoming brown to brownish (Snuff Brown, Cinnamon-brown, 5D4-6, 5E6-7) over disk, convex to plano-convex, often slightly depressed at center, viscid when moist; context white, unchanging; margin tuberculate-striate (0.27-0.4R), nonappendiculate; universal veil as conical, subconical to granular, small warts, up to 2 mm high, dirty white, grayish to yellowish (Cream Color, Pale Smoke Gray, Cream Buff, 2A1-3, 5B2), randomly arranged, frequently densely placed over disk. | ||||||||||||||||||||||||||||
| lamellae | from protolog: free, crowded, white (1A1), with finely fimbriate and floccose edges; lamellulae truncate, plentiful, in 2 - 3 ranks. | ||||||||||||||||||||||||||||
| stipe | from protolog: 40 - 90 × 5 - 10 mm, white (1A1), subcylindric or slightly tapering upward, with apex slightly expanded, fibrillose to subglabrous; context white, loosely stuffed to hollow in center; bulb 10 - 20 mm wide, white to pallid, subglobose to napiform; partial veil superior, pendent from attachment 15 - 25 mm below stipe apex, white (1A1), thin, membranous; universal veil covering upper part of bulb with floccose to granular, dirty white to yellowish or grayish (Cream Color, Pale Smoke Gray, Cream Buff, 2A1-3, 5B2) remnants. | ||||||||||||||||||||||||||||
| odor/taste | from protolog: Odor indistinct. Taste not recorded. | ||||||||||||||||||||||||||||
| pileipellis | from protolog: 100 - 150 µm thick; upper layer (60 - 80 µm thick) strongly gelatinized, comprising subradially arranged to somewhat interwoven 3 - 5 µm wide, colorless to subcolorless, moderately gelatinized, filamentous hyphae; lower layer (40 - 70 µm thick) composed of radially and compactly arranged, 3 - 6 µm wide filamentous hyphae, sometimes with intercalary segments inflated to 8 - 10 (-12) µm wide, often with brownish vacuolar pigments; vascular hyphae rare, 3 - 6 µm wide. | ||||||||||||||||||||||||||||
| pileus context | not described in protolog. | ||||||||||||||||||||||||||||
| lamella trama | from protolog: bilateral. Mediostratum 30 - 40 µm wide, composed of long ellipsoid to subfusiform, inflated cells, 90 - 130 × 10 - 30 µm; filamentous hyphae fairly abundant, 2 - 7 µm wide; vascular hyphae rare. Lateral stratum composed of long ellipsoid to fusiform inflated cells, 55 - 100 × 20 - 30 µm, diverging at an angle of 30° - 45° to the mediostratum; filamentous hyphae fairly abundant, 3 - 7 µm wide; clamps lacking. | ||||||||||||||||||||||||||||
| subhymenium | from protolog: 30 - 40 µm thick, with 2 - 3 (-4) layers of subglobose, ovoid to barrel-shaped cells, 10 - 20 × 8 - 17 µm. | ||||||||||||||||||||||||||||
| basidia | from protolog: 38 - 55 × 10 - 13 µm, clavate, 4-spored, with sterigmata 3 - 5 µm long; clamps lacking. | ||||||||||||||||||||||||||||
| universal veil | from protolog: On pileus: composed of more or less vertically (at wart base subvertically) arranged elements; inflated cells abundant to nearly dominant, subglobose, ovoid to short ellipsoid (20 - 65 × 10 - 40 µm) or ellipsoid to subfusiform (35 - 60 × 15 - 30 µm), often in chains of 2 - 4, thin-walled to slightly thick-walled, colorless, hyaline to subhyaline; vascular hyphae rare or locally conspicuous, 2 - 7 µm wide. On stipe base and upper bulb: with structure similar to that on pileus, but with elements irregularly arranged and filamentous hyphae more abundant. | ||||||||||||||||||||||||||||
| stipe context | from protolog: filamentous hyphae 2 - 7 (-10) µm wide, scattered to fairly abundant in interior, abundant on stipe surface; acrophysalides plentiful, 200 - 410 × 25 - 45 µm; vascular hyphae rare. | ||||||||||||||||||||||||||||
| partial veil | from protolog: filamentous hyphae dominating, 2 - 7 µm wide, somewhat loosely and subradially arranged, frequently branching, anastomosing, interwoven, thin-walled, colorless, hyaline; inflated cells fairly abundant to locally abundant, clavate to long ellipsoid (35 - 130 × 10 - 30 µm), occasionally subglobose to ovoid (20 - 45 × 15 - 30 µm), terminal, usually single, rarely in chains of 2 - 3; vascular hyphae rare. | ||||||||||||||||||||||||||||
| lamella edge tissue | from protolog: as sterile strip, up to 100 µm wide in side view, composed of clavate to broadly clavate to sphaeropedunculate cells (25 - 40 × 10 - 22 µm), terminal or in chains of 2 - 3, thin-walled, colorless, hyaline; filamentous hyphae very abundant, 2 - 5 µm wide, gelatinized, irregularly arranged or more or less running parallel to lamella edge. | ||||||||||||||||||||||||||||
| basidiospores |
From protolog: [360/14/7] (8.0-) 8.5 - 11.5 (-13.5) × (6.0-) 6.5 - 8.5 (-9.0) μm, ( | ||||||||||||||||||||||||||||
| ecology | Solitary to scattered. At 1200 - 2500 m elev. On soil in mixed forests including Pinus yunnanensis. | ||||||||||||||||||||||||||||
| material examined |
from protolog: CHINA: YUNNAN—Baoshan (prefecture level) City - Tengchong Co., Mingguang, 2.viii.1980 M Zang 6301 (paratype, HKAS 6301).
Chuxiong Yi Autonomous Prefecture - Wuding Co., Sishan, 27.vii.1988 L. S. Wang 9 (paratype, HKAS 20886), 10 (paratype, HKAS 20887).
Dali Bai Autonomous Prefecture - Binchuan Co., Jizushan, 9.viii.1985 G. P. Xiao 529 (paratype, HKAS 17032); Heqing Co., Huangping, 24.vii.1998 Zhu L. Yang 2490 (holotype, HKAS 32492).
Pu'er Prefecture - Simao District, Zhala Yakou, 1.viii.1991 Zhu L. Yang 1338 (HKAS 24202).
Xishuangbanna Dai Autonomous Prefecture - Menghai Co., Mangao Nature Reserve, 14.viii.1991 Zhu L. Yang 1519 (paratype, HKAS 24180). Zhang et al. (2004) vouchers for sequencing: CHINA: YUNNAN—unkn. loc., s.d.unkn. coll.s.n. (HKAS 38297), s.d.unkn. coll.s.n. (HKAS 38340), s.d.unkn. coll.s.n. (HKAS 38334). | ||||||||||||||||||||||||||||
| discussion |
from protolog: "Amanita parvipantherina is a member of Amanita [subgenus Amanita] section Amanita (Yang 1997). A few collections cited above were regarded incorrectly as A. pantherina (DC. : Fr.) Krombh., but the latter has more robust basidiocarps with shorter striations on the pileal margin, a short limbate collar on the stipe base and more abundant filamentous hyphae in the white volval remnants on the pileus (Breitenbach and Kränzlin 1995; Yang 1997, unpublished data of Yang studying material from the Netherlands [THE NETHERLANDS: ZUID-HOLLAND—Wassenaar, 2.ix.1979 C. Bas 7474 (L)]). Amanita parvipantherina is similar to A. sychnopyramis f. subannulata Hongo (Hongo 1971) [on this site treated as A. kwangsiensis Y. C. Wang—ed.], but differs from the latter by its somewhat smaller basidiocarps with a superior annulus and larger, broadly ellipsoid to ellipsoid basidiospores. The basidiospores of A. sychnopyramis f. subannulata are mostly globose to subglobose, (6.0-) 6.5 - 8.5 (-9.5) × 6.0 - 8.0 (-9.0) µm, ( | ||||||||||||||||||||||||||||
| citations | —Zhu L. Yang, M. Weiss, & F. Oberwinkler (2004) | ||||||||||||||||||||||||||||
| editors | RET | ||||||||||||||||||||||||||||
Information to support the viewer in reading the content of "technical" tabs can be found here.
| name | Amanita parvipantherina |
| bottom links | [ Keys & Checklists ] |
| name | Amanita parvipantherina |
| bottom links | [ Keys & Checklists ] |
Each spore data set is intended to comprise a set of measurements from a single specimen made by a single observer; and explanations prepared for this site talk about specimen-observer pairs associated with each data set. Combining more data into a single data set is non-optimal because it obscures observer differences (which may be valuable for instructional purposes, for example) and may obscure instances in which a single collection inadvertently contains a mixture of taxa.

Text and User-Generated Sporographs are published under the Creative Commons License.

In the case of a taxon page, image credits are on the 'image' tab.